De quaggamossel wordt door sommige waterbeheerders gezien als een zegen en zelfs ingezet om troebele wateren helder te krijgen. Monitoringsdata van de Biesboschbekkens en experimenteel onderzoek aan de onderwaterbodems, tonen echter dat een gedeeltelijk afsterven van de populatie kan leiden tot een omslag in de waterkwaliteit.
Download hier pdf van dit artikel.
De quaggamossel (Dreissena rostriformis bugensis) is een invasieve exoot die waarschijnlijk rond 2006 met ballastwater van schepen meegekomen is uit het gebied rond de Kaspische en Zwarte Zee [1]. Binnen enkele jaren heeft deze mosselsoort in veel Nederlandse meren en rivieren de driehoeksmossel (Dreissena polymorpha), een andere, reeds sinds 1800 aanwezige exoot, grotendeels verdrongen. In de Amerikaanse Grote Meren vond een soortgelijke invasie iets eerder plaats (1990). De mossel vormde hier dichte tapijten en had, mede door zijn efficiënte filtering van plankton en zwevende deeltjes, een groot effect op het voedselweb in deze meren. Er bleef minder voedsel over voor andere planktoneters, waaronder jongen van inheemse vissoorten, en het water werd glashelder, waardoor ondergedoken waterplanten zich flink konden uitbreiden.
Ook in de drie door Maaswater gevoede spaarbekkens in de Brabantse Biesbosch (De Gijster, Honderd en Dertig en Petrusplaat) komt de quaggamossel voor. Deze drie bekkens zijn de belangrijkste bron van drink- en industriewater van Evides Waterbedrijf. Maaswater wordt eerst in voorraadbekken De Gijster gepompt en stroomt vervolgens via de Honderd en Dertig naar de Petrusplaat. Daarna gaat het naar de drink- en industriewaterzuiveringen. Dankzij de bekkens is het mogelijk om periodes met een zeer lage Maasafvoer te overbruggen en bij een mindere waterkwaliteit de inname te stoppen. Het waterpeil in spaarbekken De Gijster is daardoor onderhevig aan fluctuaties terwijl dit in de andere twee bekkens normaal gesproken niet zo is. De quaggamosselpopulatie heeft zich in de Biesboschbekkens sinds 2009 sterk uitgebreid. Net als in de Amerikaanse Grote Meren is het oppervlaktewater helderder geworden en zijn er meer waterplanten gaan groeien [2]. In De Gijster heeft zich op de oorspronkelijke bodem een ’mosselbank‘ gevormd (zie afbeelding 1). De dichtheid van levende mosselen bedroeg in 2014 gemiddeld 3000 per m2 en de biomassa aan mosselvlees bedroeg 25 g per m2 [3].
Afbeelding 1. Drooggevallen mosselbank in spaarbekken De Gijster (2015)
Mosselsterfte
De vraag is of de mosselen niet ook een negatief effect kunnen hebben op de waterkwaliteit, met name wanneer een deel van de mosselpopulatie in korte tijd afsterft. Dit is een reële mogelijkheid bij een langdurige innamestop als gevolg van een langdurige verontreiniging van het Maaswater. In theorie kan een slechte waterkwaliteit direct leiden tot mosselsterfte. Deze kan echter ook indirect bijdragen aan mosselsterfte omdat bij een slechte waterkwaliteit geen water ingenomen wordt in De Gijster, waardoor het waterpeil zakt en mosselbanken deels droogvallen en afsterven. Dit kan ook weer gevolgen hebben voor de mosselbanken onder water. Er is vastgesteld dat het gehalte aan DOC (opgelost organisch koolstof), ammonium, nitraat en fosfor in de waterkolom van De Gijster na het droogvallen van de bodem (2015) en het weer opvullen van het bekken inderdaad toenam [4]. Ook kan een sterk lagere fytoplanktonproductie leiden tot mosselsterfte als gevolg van voedselgebrek. Daarnaast is het bekend dat gedurende de warme zomerperioden in bepaalde delen van de bekkens nabij de bodem zuurstofloosheid kan optreden, met mogelijke mosselsterfte tot gevolg. Om meer inzicht te krijgen in de mate waarin (interacties tussen) bodemprocessen en quaggamosselen de waterkwaliteit kunnen beïnvloeden heeft onderzoekscentrum B-WARE onderzoek uitgevoerd met behulp van een groot aantal intacte bodemprofielen (20-30cm) afkomstig uit de drie Biesboschbekkens.
Onderzoek
In maart 2016 zijn in De Gijster op vijf locaties mosselen verzameld in potten (20x20x30cm). Hiervan werden de mosseldichtheid en mossellengten met bijbehorende vers- en drooggewichten van de schelp en weke delen, alsmede de chemische samenstellingen van de schelp en weke delen bepaald. De dikte van de mosselbank bedroeg gemiddeld 16,2 cm (12 tot 20 cm). In september 2016 zijn, verspreid over de drie bekkens, op 31 locaties in duplo op een diepte van 5 tot 27 meter profielen van de bodem (20-30 cm) gestoken met behulp van een uwitec-sampler voorzien van hamerfunctie. De kolommen hadden een hoogte van 60 cm en een diameter van 5,5 cm.
Het effect van de mosseldichtheid op de waterkwaliteit is onderzocht onder zuurstofhoudende en zuurstofloze condities in een donkere klimaatcel bij een constante temperatuur van 18°C. Daarbij werd oppervlaktewater uit de drie bekkens gebruikt. Zuurstofloze condities werden opgewekt door het oppervlaktewater te doorborrelen met stikstofgas, alvorens dit in de kolommen aan te brengen en de waterlaag af te sluiten met een dun laagje paraffineolie (afbeelding 2). Op T=0, T=3, T=7, T=14 en T=56 dagen werden oppervlaktewatermonsters verzameld. Poriewatermonsters werden verzameld op T=0. Op het einde van het experiment werd per cilinder het aantal mosselen geteld en de toplaag van de bodem (0-2 cm) verzameld voor chemische analyses.

Afbeelding 2. In sterk reductieve bodems en/of in een zuurstofarme waterlaag (rechts) worden in de bodemtoplaag en op de schelp zwarte ijzersulfiden gevormd (mosselsterfte) (kolom uit Petrusplaat)
Anaerobe afbraakprocessen in de bodem van de Biesboschbekkens
De mosselbank bestaat uit levende mosselen met daaronder de resten van gestorven mosselen en (pseudo)feces. Deze laag bevat circa 3,3 kg organisch materiaal per m2, waarvan 49,2 gram per m2 uit fosfor (P) bestaat. Bij de afbraak van organisch materiaal komt anorganisch koolstof vrij in de vorm van kooldioxide en bicarbonaat (CO2 en HCO3-). Het totale anorganisch koolstofgehalte (total inorganic carbon, TIC) is daarom een goede maat voor de mate waarin er in de onderwaterbodem (anaerobe) afbraak van organisch materiaal plaatsvindt.
Geoxideerd ijzer en sulfaat (SO42-) kunnen (bij de afwezigheid van zuurstof) dienen als alternatieve elektronenacceptor voor de afbraak van organisch materiaal. Relatief goed oplosbaar tweewaardig ijzer (Fe2+) wordt daarbij vrijgemaakt uit de reductie van slecht oplosbaar driewaardig ijzer (ijzer(III)(hydr)oxides). Hierbij wordt niet alleen ijzer vrijgemaakt maar komt ook fosfor vrij dat geadsorbeerd is aan de ijzer(III)(hydr)oxides. Bij de reductie van sulfaat wordt vrij sulfide gevormd dat neerslaat met opgelost tweewaardig ijzer tot ijzersulfide (zwart). Hierdoor vindt er in de bodem geen accumulatie plaats van het potentieel giftige vrije sulfide (S2-) en krijgt de bodem een zwarte kleur (zie afbeelding 2). Uit afbeelding 3 blijkt dat in de onderwaterbodems uit de Biesboschbekkens de ijzer-, ammonium- en fosforconcentratie in het poriewater positief correleerden met de TIC-concentratie, terwijl de sulfaatconcentratie negatief correleerde met de TIC-concentratie. Dit bewijst dat bovengenoemde processen een grote rol spelen in de mobilisatie van nutriënten in onderwaterbodems van de Biesboschbekkens. Vastgesteld is dat na het droogvallen van de bodem en het weer opvullen, het gehalte aan DOC, ammonium, nitraat en fosfor in de waterkolom inderdaad toenam [3]. Het sulfaat wordt daarbij aangevoerd via het Maaswater.
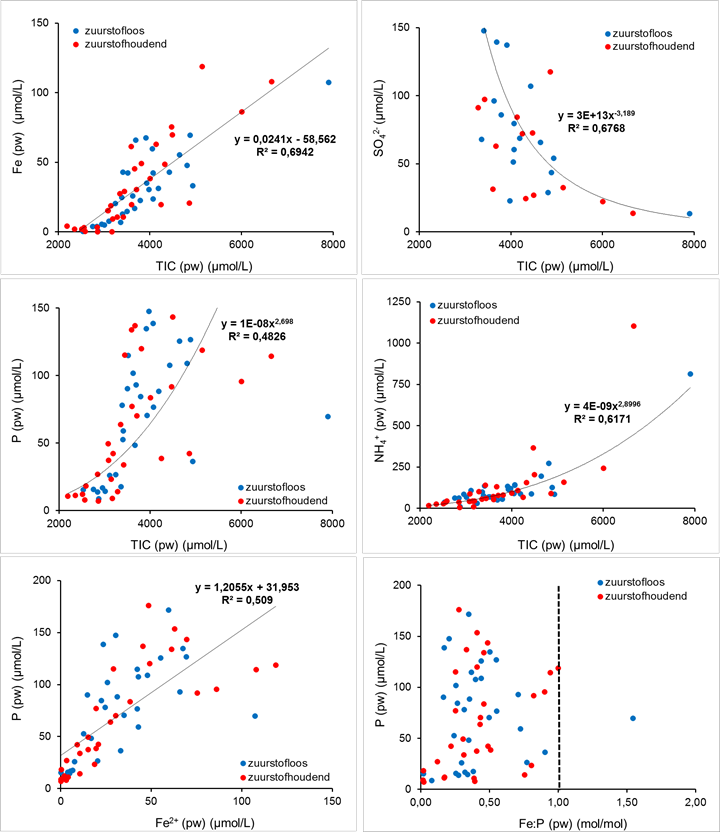
Afbeelding 3. Correlaties in het poriewater (pw) laten zien dat anaerobe afbraakprocessen plaatsvinden in de onderwaterbodems van de Biesboschbekkens, onafhankelijk van het zuurstofgehalte in de waterlaag. De zwarte stippellijn geeft de ratio aan waaronder fosfaatmobilisatie naar de waterlaag plaats kan vinden onder aerobe omstandigheden
P-nalevering vanuit de bodem
Voor de meeste onderwaterbodems uit de Biesboschbekkens was de fosfaatnalevering laag onder aerobe omstandigheden en veel hoger onder anaerobe condities (afbeelding 4). Onder aerobe omstandigheden wordt op de overgang van de waterbodem naar de waterlaag opgelost gereduceerd ijzer (Fe2+) geoxideerd tot ijzer(III)(hydr)oxiden. Aan de gevormde ijzer(hydr)oxiden wordt fosfaat goed gebonden (de ijzerval). Daarnaast kan zuurstof in de bodem ook ijzersulfide oxideren. Hierbij wordt sulfaat weer teruggevormd en komt Fe2+ vrij, dat in de toplaag weer kan worden geoxideerd tot Fe3+. Onder anaerobe omstandigheden wordt geen ijzer geoxideerd, waardoor fosfaat (en ijzer) vrij de waterlaag in kunnen diffunderen. Het is bekend dat de nalevering van P naar de zuurstofhoudende waterlaag sterk toeneemt wanneer de Fe:P-ratio in het poriewater lager wordt dan 1 [4]. Ook ammonium kan uit de onderwaterbodem naar de waterlaag diffunderen en hier accumuleren, wat ook werd vastgesteld bij andere experimenten waarbij bodemmateriaal van de bekkens werd geïncubeerd [5].
Sommige bodems uit de Biesboschbekkens, met een van origine zuurstofhoudende waterlaag, leverden toch fosfaat na. Deels wordt dit veroorzaakt door de relatief lage Fe/P-ratio’s van het poriewater. Daarnaast is de waterlaag in een aantal van deze kolommen toch anaeroob geworden. Deze waren tijdens het experiment goed herkenbaar omdat de toplaag van de bodem, net als in de anaerobe kolommen, ook zwart kleurde (zie afbeelding 2). Deze kolommen kwamen van locaties waar de waterlaag nabij de bodem zuurstofloos wordt gedurende de zomer. De betreffende kolommen zijn in afbeelding 4 gemarkeerd met een kruisje. Het gaat hierbij in alle gevallen om kolommen met mosselen. Deze kolommen onderscheidden zich ook omdat er in de waterlaag ammonium accumuleerde. Het is niet duidelijk of deze kolommen anaeroob werden vanwege het zuurstofgebruik door de mosselen en/of de onderwaterbodem.
In de kolommen met een zuurstofloze waterlaag was gemiddeld 4,5 mg fosfor per vierkante meter aanwezig in de weke mosselbiomassa en 193 mg fosfor per vierkante meter in de schelpbiomassa. Bij het afsterven van de mosselen kan het fosfor uit de weke mosselbiomassa vrijkomen. In de anaerobe kolommen met mosselen kwam gedurende het experiment echter gemiddeld 75 maal zoveel P vrij als in de weke mosselbiomassa aanwezig was. Dit betekent dat de nalevering van fosfor niet kan worden verklaard door het vrijkomen van fosfor uit de dode mosselen en dat het nageleverde fosfor vooral afkomstig was uit de onderwaterbodem. Dit verklaart waarom de nalevering van fosfor naar de waterlaag bij de anaerobe incubaties gecorreleerd was met de fosforconcentratie in het poriewater van de onderwaterbodem en minder met de mosseldichtheid. Sinds de mosselen zich hebben gevestigd bestaat de toplaag van de onderwaterbodem in de Biesboschbekkens uit lege mosselschelpen van afgestorven schelpen (de onderste mosselen sterven wanneer nieuwe dieren zich op deze mosselen vestigen) met daartussen opgehoopt feces en pseudofeces rijk aan organisch materiaal. Metingen in 2016 toonden dat in De Gijster per m2 ongeveer 3,3 kg organische stof, 182 kg schelpmateriaal en 49,2 gram P aanwezig was [6].
Afbeelding 4. De nalevering van fosfaat en ammonium onder zuurstofloze en zuurstofhoudende condities uitgezet tegen de concentraties in het poriewater (pw, links) en de mosseldichtheid (rechts). De rode stippen met kruis representeren locaties met mosselen waar de waterlaag door respiratie zuurstofloos wordt
N-nalevering vanuit de bodem
De nalevering van ammonium liet, in tegenstelling tot die van fosfaat, geen verband zien met de ammoniumconcentratie in het poriewater, maar bleek sterk gecorreleerd met het aantal mosselen in de kolom (R2= 0,8895). Het afsterven van mosselen in de zuurstofloze waterlaag leidde duidelijk tot het vrijkomen van ammonium. Ook in kolommen, waarin de waterlaag gedurende het experiment zuurstofloos is geworden, was de ammoniumconcentratie gecorreleerd met het aantal mosselen in de kolom. Alleen voor de kolommen met een zuurstofloze waterlaag en met weinig of geen mosselen was er een correlatie met de ammoniumconcentratie in het poriewater.
Ook in de kolommen met een zuurstofloze waterlaag, zonder mosselen en met een zeer hoge ammoniumconcentratie in het poriewater werd een verhoogde ammoniumnalevering naar de waterlaag gemeten. Ammonium wordt in de waterlaag door nitrificerende bacteriën geoxideerd tot nitraat (NO3-) dat vervolgens in de anaerobe onderwaterbodem wordt gedenitrificeerd tot stikstofgas (N2). Hierdoor vindt er geen ophoping plaats van ammonium in de kolommen met zuurstofhoudend water.
Voor stikstof geldt dat er voor de anaerobe kolommen gemiddeld 3.840 mg N per vierkante meter aanwezig was in de weke schelpbiomassa en 3.900 mg N per vierkante meter in de schelpbiomassa. Tijdens het twee maanden durende naleveringsexperiment kwam er in de anaerobe kolommen met schelpen gemiddeld 32% van alle stikstof in de weke schelpbiomassa vrij als ammonium in de waterkolom. Dit laat zien dat er bij het afsterven van de mosselen gedurende enkele maanden een zeer sterke nalevering van stikstof naar de waterlaag kan zijn als gevolg van het afsterven van de mosselen. De N-nalevering (mg-N/m2/dag) is gelijk aan 0,0054x + 3,3846, waarbij x= het aantal mosselen per m2 (afbeelding 4, rechtsonder).
Conceptueel model
Dreissena-mosselen kunnen bij een temperatuur van 20°C enkele dagen tot een week overleven onder zuurstofloze omstandigheden of bij droogval [7]. Uit ons experiment is gebleken dat de waterlaag op locaties met veel quaggamosselen zuurstofloos kan worden door afbraakprocessen in de onderwaterbodem. Respiratie door mosselen en afbraak van organisch materiaal van (pseudo)feces zal dit bespoedigen. In de veldsituatie is de waterkolom veel hoger, waardoor er microstratificatie boven het sediment kan optreden en de waterlaag boven de onderwaterbodem sneller anaeroob kan worden. Zuurstofloze condities treden in De Gijster ook daadwerkelijk op gedurende de zomer [2].
Onder zuurstofloze omstandigheden werkt de ijzerval niet meer en wordt de nalevering van fosfor naar de waterlaag met name gestuurd door de fosfaatconcentratie in het poriewater van de onderwaterbodem [5]. De aanwezigheid van quaggamosselen kan leiden tot een hogere P-nalevering, maar het fosfor dat vrijkomt is maar voor een klein deel afkomstig uit de afstervende mosselen (2,3 % van het totale P) en voor het overgrote deel uit de onderliggende laag met resten van mosselen en mosselfeces. De nalevering van ammonium onder anaerobe condities wordt vrijwel volledig bepaald door de mosseldichtheid. Ammonium wordt uitgescheiden door levende mosselen, maar komt in sterk verhoogde concentraties vrij bij het afsterven van de mosselen.
De aanwezigheid van quaggamosselen kan zorgen voor helder water omdat de mosselen algen uit het water filtreren en daarbij een relatief grote hoeveelheid (pseudo)feces produceren die zich tussen de schelpresten van afgestorven mosselen ophogen.
Omslag waterkwaliteit
Wanneer in een bekken een relatief grote bedekking aanwezig is van quaggamosselen, kan het (deels) instorten van de populatie leiden tot een zeer plotselinge omslag van de waterkwaliteit. Wanneer er een aanzienlijke sterfte optreedt, door zuurstofgebrek of wat voor andere reden dan ook, kan als gevolg de zuurstofconcentratie (lokaal) sterk dalen, waardoor de anaerobe zone nabij de bodem zich verder kan uitbreiden, waardoor nog meer sterfte optreedt. Tegelijkertijd neemt de nalevering van fosfor (uit de onderwaterbodem, met pseudofaeces) en ammonium (m.n. uit de afstervende mosselen) sterk toe. Dit kan leiden tot een explosieve groei van algen omdat er ook geen algen meer door de mosselen worden gefilterd uit de waterlaag. Hierdoor wordt het water troebel waardoor ook de aanwezige waterplantenvegetaties kunnen afsterven, waarbij ook weer nutriënten vrijkomen.
Massale sterfte van Dreissena-mosselen kan optreden bij een verlaging van de pH-waarde en de calciumconcentratie van het water (< 8,5 mg Ca/L), bij het ontstaan van zuurstofloze condities door organische belasting en hogere temperaturen, als gevolg van droogval bijvoorbeeld bij een lage rivierafvoer gedurende strenge winters [8], [9]. Indien deze of andere scenario’s (bijvoorbeeld infecties) plaatsvinden kan er dus gedurende een periode van maanden aanzienlijke stikstof- maar ook fosfornalevering plaatsvinden naar de waterlaag. De stikstofnalevering wordt dan volledig bepaald door de mosseldichtheid. Een schematisch overzicht van het conceptueel model van de relatie van de quaggamossel met de waterkwaliteit is gegeven in afbeelding 5.
Het onderzoek heeft dus aangetoond dat systemen die helder zijn geworden door de aanwezigheid van quaggamosselen dus ook erg kwetsbaar zijn als de mosselbanken (deels) afsterven.

Afbeelding 5. Conceptueel model van de relatie van de quaggamossel met de waterkwaliteit. Een stippellijn geeft een remming weer
Referenties
- Matthews, J. et al. (2014). Rapid range expansion of the invasive quagga mussel in relation to zebra mussel presence in The Netherlands and Western Europe. Biological Invasions 16(1): 23–42.
- Wagenvoort, A. (2014). Memo Evaluatie 10 jaar Biesboschspaarbekkens: veldparameters, voedingsstoffen en algenbiomassa. Wilhelminadorp, 20 juni 2014, 6 pp.
- Wagenvoort, A. (2015). Quaggamosselen in de Biesboschspaarbekkens - Overzicht van een uitgebreide inventarisatie najaar 2014. Rapport. AqWa-ecologisch advies, Wilhelminadorp, 62 pp.
- Geurts, J.J.M. et al. (2010). The interaction between decomposition, N and P mineralization and their mobilization to the surface water in fens. Water Research 44: 3487-3495.
- Wagenvoort, A. & Schaaf, B. (2016). Effecten van een langdurige innamestop op de waterkwaliteit. Rapport. AqWa ecologisch advies/Evides waterbedrijf Rotterdam, 26 pp.
- Stelten, F.J.A. (2016). Indicatief waterbodemonderzoek Spaarbekken "De Gijster" in de Biesbosch. Rapport BK Ingenieurs, Dordrecht, 73 pp.
- Claudi, R. & Mackie, G.L. (1994). Practical manual for zebra mussel monitoring and control. Lewis Publishers, CRC Press, Boca Raton, Florida.
- Hinks, S. S. & Mackie, G. L. (1997). Effects of pH, calcium, alkalinity, hardness, and chlorophyll on survival, growth, and reproductive success of zebra mussel (Dreissena polymorpha) in Ontario lakes. Canadian Journal of Fisheries and Aquatic Sciences, 54: 2049-2057.
- Leuven, R.S.E.W. et al. (2014). Mass mortality of invasive zebra and quagga mussels by desiccation during severe winter conditions. Aquatic Invasions 9/3: 243-252.















